- 品牌
- 上海保藏生物技术中心 SHBCC
- 贮藏
- 4-10度冷藏
- 生产企业
- 瑞楚生物科技(江苏)有限公司
- 规格
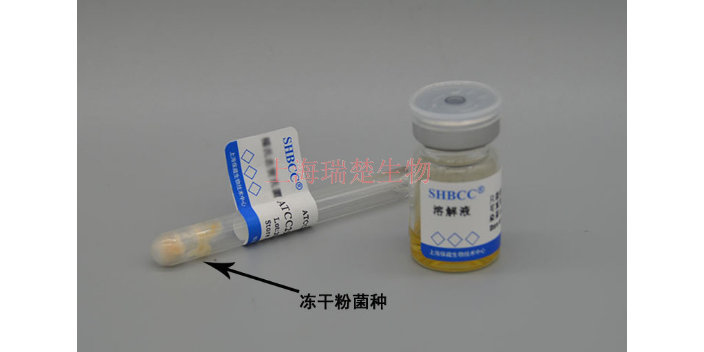
- 冻干管
- 产地
- 江苏盐城
- 厂家
- 上海保藏生物技术中心
- 有效期
- 36个月
- 运输条件
- 冰袋加顺丰快递
- 长期储存温度
- 4-10度
- 活化之后储存条件
- 4-10度
- 转接时间
- 3个月转接一次
- 长期储存方法
- 甘油菌-80度可以保存2年以上
- 活化方法
- 好氧菌斜面活化,厌氧菌培养皿厌氧培养,兼性厌氧液体培养基培养
- 打管方法
- 直接敲开
- 接种量
- 活化时取0.2-0.3ml溶解液后全部溶解接种到培养基上
- 活化代数
- 活化3代活力比较好
- 操作硬件设施要求
- 二级生物安全柜内操作
棉花新鞘氨醇菌(Novosphingobiumgossypii)在生物修复领域具有一些潜在的应用,尽管搜索结果中没有直接详细描述其具体的应用案例。然而,基于其所属的Novosphingobium属的特性,可以推测其在以下方面可能具有应用潜力:1.**降解有机污染物**:Novosphingobium属的细菌普遍具有降解芳烃(芳香族)化合物的特性,是良好的芳烃污染环境的生物修复菌。棉花新鞘氨醇菌可能也具有类似的降解能力,能够分解环境中的有机污染物。2.**趋化性研究**:研究表明,新鞘氨醇杆菌对芳香族化合物和TCA循环中间代谢物具有不同程度的趋化性。这种趋化性可能有助于细菌在污染环境中寻找并降解污染物,从而在生物修复中发挥作用。3.**环境适应性**:棉花新鞘氨醇菌的革兰氏阴性杆菌特性和不产芽胞的特点,使其在不同环境条件下具有一定的生存能力。这种适应性可能有助于其在复杂环境中进行生物修复。4.**基因组研究**:通过对棉花新鞘氨醇菌的基因组研究,可以揭示其降解污染物的代谢途径和调控机制。这有助于开发更有效的生物修复策略。5.**生态修复**:棉花新鞘氨醇菌可能在生态修复中发挥作用,特别是在处理土壤和水体中的有机污染物时。其降解能力可以帮助恢复受污染环境的生态平衡。橙色螺状菌具有生物活性物质合成的潜力,能够产生胞外酶,分解纤维素、琼脂、几丁质等 。蜡蚧菌

棉花新鞘氨醇菌(Novosphingobiumgossypii)是一种属于Novosphingobium属的微生物,具有一些独特的特征和应用价值。以下是关于棉花新鞘氨醇菌的一些关键信息:1.**形态特征**:棉花新鞘氨醇菌是一种革兰氏阴性杆菌,它不产芽胞并且能够产生黄色素。它的主要醌型为Q-10,主要脂肪酸包括C18:1ω7c(占70%)和2-羟基脂肪酸,C14:02-OH。2.**原产地**:这种微生物的原产地为美国,具体是从阿拉巴马州塔拉西的健康陆地棉品种DES-119的茎组织内部分离得到的。3.**主要用途**:棉花新鞘氨醇菌的主要用途是进行分类学研究。它的16SrRNA基因序列号为KP657488。4.**培养条件**:关于具体的培养条件和培养基,搜索结果中没有提供详细信息。但是,一般的培养方法包括准备含预除氧液体培养基的试管,在安全柜中将菌粉溶解后再置于相应培养条件下。5.**保存与使用**:在使用棉花新鞘氨醇菌时,需要注意活化前将冷冻管置于低温、干燥处,避免菌种衰退。开封、复溶等操作应无菌进行。如发现冷冻管盖松、复溶液浑等异常,应停止使用。保存时根据细菌特性选择合适的培养基,并注意不同细菌的保存温度。定期转种,每3代鉴定一次。唐德链霉菌菌株居海绵华美菌属于细菌界,由韩国济州国立大学的B.-J. Yoon分离并命名 。
拉氏根瘤菌(Rhizobiumleguminosarum)主要与豆科(Fabaceae)植物形成共生固氮关系,其作用机制在其他类型的植物中并不相同。以下是一些原因和差异:1.**宿主专一性**:拉氏根瘤菌对豆科植物具有高度的宿主专一性,它们的Nod因子和其他共生信号分子专门针对豆科植物的识别系统。2.**不同植物家族的根瘤菌**:不同植物家族有不同的根瘤菌与之共生。例如,苜蓿科(Fabaceae)植物通常与慢生根瘤菌(Bradyrhizobium)共生,而其他非豆科植物则可能不形成根瘤或与不同类型的固氮菌共生。3.**共生信号的差异**:不同植物家族释放的信号分子和根瘤菌产生的Nod因子在结构和功能上可能有所不同,导致它们之间的共生信号交流机制存在差异。4.**根瘤结构的不同**:即使在能够形成根瘤的植物中,根瘤的结构和发育过程也可能因植物种类而异。例如,一些植物可能形成簇状根瘤,而另一些则形成单个根瘤。5.**固氮酶系统的适应性**:拉氏根瘤菌的固氮酶系统适应于豆科植物的共生固氮需求,可能不适应其他植物的生理和代谢特性。6.**基因表达和调控的差异**:在与非豆科植物相互作用时,拉氏根瘤菌可能无法正确表达或调控其共生基因,导致无法形成有效的共生关系。
棉花新鞘氨醇菌(Novosphingobiumgossypii)作为一种新鞘氨醇菌属的细菌,可能具有以下生物修复中的降解机制,尽管具体的机制可能需要通过实验室研究来明确:1.**芳香族化合物的降解**:新鞘氨醇菌属的细菌通常具有降解芳香族化合物的能力。棉花新鞘氨醇菌可能通过其代谢途径中的酶系统,将芳香族化合物转化为中间代谢产物,后完全矿化为二氧化碳和水。2.**电子传递链**:在降解过程中,棉花新鞘氨醇菌可能利用其电子传递链中的酶,如加氧酶和脱氢酶,将有机污染物氧化,生成更易降解的化合物。3.**共代谢途径**:该菌可能通过共代谢途径参与污染物的降解,即在降解其自身生长所需的营养物质的同时,也对环境中的污染物进行转化。4.**酶促反应**:棉花新鞘氨醇菌可能产生特定的酶,如漆酶、过氧化物酶、或者特定的加氧酶,这些酶能够催化有机污染物的降解反应。5.**基因表达调控**:在生物修复过程中,细菌可能会根据环境条件调节其基因表达,以适应污染物的降解需求。棉花新鞘氨醇菌可能具有这样的调控机制,以优化其降解途径。6.**适应性进化**:长期暴露在污染物中可能促使棉花新鞘氨醇菌发生适应性进化,增强其降解特定污染物的能力。居海绵华美菌的培养条件为DSMMedium 514,在28°C的条件下培养,可以使用BACTO MARINE BROTH 作为培养基 。
巴塞尔贪铜菌(Cupriavidusbasilensis)是一种属于β-变形菌纲的革兰氏阴性细菌。以下是一些关于巴塞尔贪铜菌的特点:1.**代谢多样性**:巴塞尔贪铜菌能够利用多种碳源进行生长,表现出代谢多样性。2.**耐重金属**:这种细菌对重金属如铜(Cu)具有较高的耐受性,能够在含有重金属的环境中生存。3.**植物生长促进**:巴塞尔贪铜菌与某些植物共生,能够促进植物生长,可能与植物根系形成共生关系。4.**生物修复潜力**:由于其耐重金属的特性,巴塞尔贪铜菌在生物修复领域具有潜在的应用价值,特别是在重金属污染的土壤和水体的修复中。5.**环境分布**:巴塞尔贪铜菌在自然环境中分布广,可以在土壤、水体和沉积物等多种环境中找到。6.**遗传特性**:巴塞尔贪铜菌的基因组包含多种与重金属耐受性和代谢途径相关的基因,这些基因为其提供了环境适应性。7.**生理生化特性**:巴塞尔贪铜菌的生理生化特性包括其呼吸类型、酶活性和代谢途径,这些特性有助于其在不同环境条件下的生存和代谢活动。8.**分类学地位**:在分类学上,巴塞尔贪铜菌属于Ralstonia属,该属的细菌在环境微生物学和生物技术领域中具有重要的地位。稍白长孢菌(Longispora sp.)是一种革兰氏阳性细菌,其形态特征为不游动、不抗酸,具有分枝的基丝。根瘤菌
居海绵华美菌,这种细菌是从大韩民国济州沿海地区的海洋海绵中分离出来的。蜡蚧菌
巴塞尔贪铜菌(Cupriavidusbasilensis)在生物修复重金属污染方面具有潜在的应用,尽管搜索结果中没有直接提及该菌种具体的生物修复机制。然而,基于其特性和β-变形菌纲的一些共性,我们可以推断其可能的生物修复机制:1.**重金属耐受性**:巴塞尔贪铜菌可能具有耐受多种重金属的能力,这使得它能够在重金属污染的环境中生存并发挥作用。2.**生物吸附**:该菌可能通过细胞表面的官能团与重金属离子形成稳定的复合物,从而吸收和固定重金属离子,减少其在环境中的迁移性和生物有效性。3.**生物转化**:巴塞尔贪铜菌可能具有将重金属转化为较低毒性形态的能力,例如将六价铬还原为三价铬,或将有机砷氧化为无机砷等。4.**植物-微生物联合修复**:巴塞尔贪铜菌可能与超积累植物形成共生关系,通过分泌植物生长调节物质和有机配位体,促进植物对重金属的吸收和转运,提高植物修复的效率。5.**分泌有机酸**:该菌可能通过分泌有机酸(如柠檬酸、琥珀酸等)与重金属离子络合,改变土壤中重金属的存在形态,降低其毒性并促进植物吸收。6.**产生铁载体**:巴塞尔贪铜菌可能通过产生铁载体与重金属离子络合,减少宿主植物对重金属的吸收,从而减轻重金属对植物的危害作用。蜡蚧菌