4.脂质体的性质:脂质体的形态、大小、表面电荷等性质会影响药物的载药率。例如,小尺寸的脂质体通常具有较高的表面积,有利于药物的扩散和溶解。5.药物与脂质体的相互作用:药物与脂质体之间的相互作用形式也会影响载药率,例如药物与脂质质体之间的静电相互作用、疏水相互作用等。评估脂质体的载药率通常需要进行药物释放实验或者溶解度测定等试验,以确定药物在脂质体中的含量或者释放速率。通过优化脂质体的组成和制备方法,可以提高脂质体的载药率,从而增强其在药物传递等应用中的效果。脂质体制备方法:薄膜⽔化法。四川脂质体载药动物实验

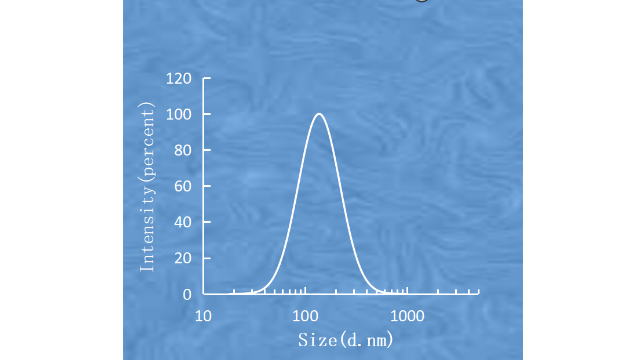
脂质体的粒径和粒径分布脂质体的整个药代动⼒学过程,如全⾝循环和MPS***、外渗到组织间质、细胞外基质间质运输以及细胞摄取和细胞内运输,都是依赖于尺⼨的。粒径<200nm的颗粒可降低⾎清蛋⽩的调理作⽤,降低MPS的***率。在⼩⿏⽩⾎病模型中,对于Myocet来说,较⼩的脂质体具有更⾼的抗**功效和增加的平均⽣存时间。粒径为2.0-3.5µm的Mepact可促使单核细胞/巨噬细胞吞噬,触发*****的免疫调节作⽤。Singh等⼈发现,含有不同颗粒⼤⼩的佐剂脂质体(ArmyLiposomeFormulation,ALF)的疫苗会产⽣不同的免疫反应,即树突状细胞更有效地摄取10-200nm范围内的⼩颗粒,⽽其他免疫细胞,如巨噬细胞,则倾向于吞噬⼤颗粒。Niu等⼈研究了⼝服给药的胰岛素负载脂质体,发现直径为150nm和400nm的脂质体表现出较慢且持续时间⻓达24⼩时的降糖作⽤,⽽粒径约为80nm和2µm的脂质体则分别表现出短暂且⽆药理作⽤。文献表明,对于*****的脂质体来说,小于200nm的脂质囊泡大小可以从物理肝脏筛选过程中逃逸。根据肝窦的大小,需要小于150nm的囊泡才能通过高渗透性的**血管穿透到恶性组织中。因此,它是由增强的渗透率(EPR)效应控制的,这有助于脂质体通过被动靶向在**中积累。上海济南脂质体载药质粒DNA要在细胞内被有效地翻译,质粒DNA必须经过有效的细胞内运输进入细胞质,并从细胞质进入细胞核。
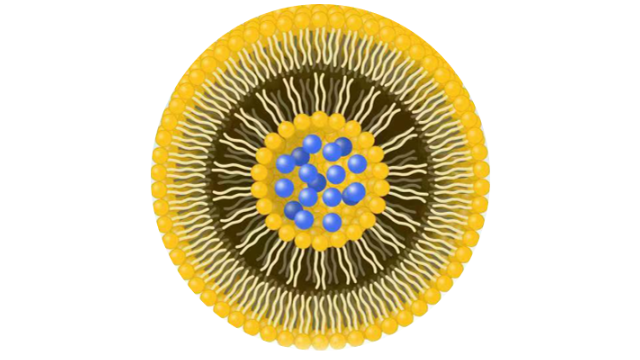
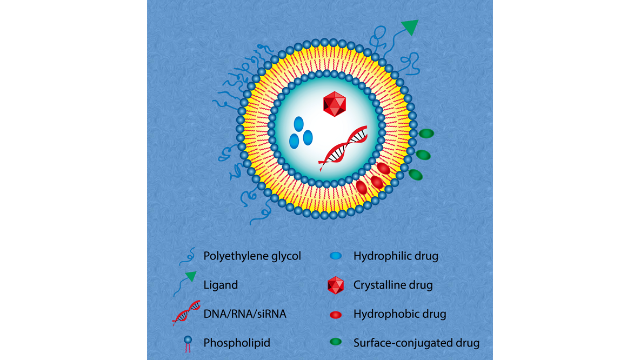
脂质体成分配比脂质体是由多种组分构成的,
主要包括:1.磷脂质:是脂质体**主要的组分,构成了脂质双层结构的主体。磷脂质包括磷脂、甘油磷脂、胆固醇等,它们在水性环境中通过亲水头部和疏水尾部的相互作用形成了双层结构。2.胆固醇:在脂质体中扮演着调节脂质双层流动性和稳定性的重要角色。胆固醇可以调节磷脂质的包装密度,增强脂质体的机械稳定性。3.表面活性剂:通常用于稳定脂质体的水合壳,并且有助于脂质体的稳定分散在水相中。常见的表面活性剂包括辛酸单酯类、磺酸盐类等。4.PEG衍生物:如前面所述,聚乙二醇(PEG)衍生物可以修饰脂质体表面,增强其稳定性、延长血液循环时间和降低免疫原性。5.药物或其他活性成分:脂质体通常被设计用来载药或其他活性成分,这些物质可以被包裹在脂质体内部,通过脂质双层的特性来实现针对性的释放或传递。DepoCyte、DepoDur和Exparel具有特殊的结构和相似的脂质成分。MVLs的形成⾄少需要两种类型的脂质:两亲性脂质和中性脂质(如双⽢油酯、⽢油三酯、植物油)。
与化学增敏剂共同递送为了增强***活性,研究人员研究了将***siRNA和化学药物共同装载到阳离子脂质体中的共递送方法。例如,将丝裂原活化的蛋白激酶抑制剂PD0325901包封在由N、N-二油基谷酰胺阳离子脂质、DOPE和胆固醇组成的阳离子脂质体中,通过静电相互作用与Mcl-1siRNA络合。在小鼠模型中,瘤内给药这些阳离子脂质体可***抑制**生长。在另一项研究中,开发了基于三叶赖氨酸油酰酰胺的阳离子脂质体,用于共同递送Mcl-1siRNA和***药物亚酰苯胺羟肟酸。与Mcl-1siRNA脂质体或含亚甲基苯胺羟肟酸脂质体的单药***相比,使用载药聚乙二醇化脂质体与Mcl-1siRNA复合物可提高荷瘤小鼠的体内***效果。***,将多柔比星包裹的阳离子脂质体与编码磷酸化缺陷小鼠survivin蛋白的质粒DNA复合,该蛋白是BIRC5基因编码的一种致*蛋白,是凋亡抑制剂家族的成员,苏氨酸34-丙氨酸突变体,然后用缩短的人碱性成纤维细胞生长因子肽修饰,对表达成纤维细胞生长因子受体的细胞产生选择性。在静脉给药这些复合物后,在患有肺*的C57BL/6小鼠中观察到**生长的***降低。目前临床应用面临的挑战。聚乙二醇在免疫脂质体中起到了重要作用。
脂质体靶向递送中RGD配体修饰尽管阳离子脂质体具有在体内递送核酸的潜力,但其递送到特定靶点仍然是一个主要挑战。为了增强携带核酸的阳离子脂质体在靶组织中的分布,研究人员用多肽和小分子修饰了脂质体表面。例如,研究了Arg-Gly-Asp(RGD)肽修饰的脂质体增强核酸向整合素受体表达细胞传递的能力。负载P糖蛋白特异性siRNA的RGD修饰阳离子脂质体对整合素受体表达的人乳腺*MCF7/A细胞的递送率更高,导致P糖蛋白的***沉默。与此一致的是,分子成像显示,与小鼠模型的邻近正常组织相比,MCF7/A**组织中RGD修饰的阳离子脂质体和siRNA的分布更高。在**近的一项研究中,用环RGD和辛精氨酸修饰脂质体表面,以利用环RGD的整合素受体结合效应和辛精氨酸的细胞穿透效应。双配体修饰的阳离子脂质体增加了整合素avb3表达细胞的细胞摄取,并且更有效地转染荧光素酶编码质粒DNA。Zeta电位被认为是影响细胞摄取和药物传递的重要因素之一。青海脂质体载药动物实验
阳离子脂质体提高siRNA的细胞递送和基因沉默效率。四川脂质体载药动物实验
脂质体靶向递送中叶酸配体修饰脂质与生物活性小分子(如叶酸)的结合已被研究用于靶向递送核酸。例如,由叶酸与1-棕榈酰-2-油酰-sn-甘油-3-非共价结合而形成的脂质体乙基磷脂胆碱:胆固醇脂质体显著提高胸苷激酶质粒DNA转染效率,抑制体外TSA和SCC7细胞生长。这些叶酸相关的脂质体在移植SCC7**的小鼠中显示出较高的抗**效果。在另一种方法中,叶酸标记的阳离子脂质体与小牛胸腺DNA复合物***巨噬细胞,与不含叶酸的普通阳离子脂质体相比,显示出更高的DNA叶酸受体表达细胞的递送。在荷瘤小鼠中,与不含叶酸的脂质体相比,叶酸标记的脂质体诱导干扰素-g和白细胞介素-6的产生,延长了存活时间。甘草次酸已被用于靶向肝细胞肝*细胞,基于一项研究表明,与邻近的非**肝细胞相比,甘草次酸的结合靶点蛋白激酶C在肝细胞*细胞表面的表达更高。合成了甘次酸-次酸-聚乙二醇-聚胆甾醇缀合物,并将其与DOTAP和胆固醇配制成阳离子脂质体。这些脂质体与表达GFP的质粒DNA形成复合物的能力更高,并且与缺乏甘次酸的对照阳离子Lipo脂质体相比,能增强质粒DNA转染至肝*细胞的能力。四川脂质体载药动物实验